
Variation of the linking loop in dimeric α-β barrel proteins
SUPERVISOR: Stefan HOFBAUER
Background.
Chlorite dismutases (Clds), coproheme decarboxylases (ChdCs) and dye-decolorizing peroxidases (DyPs) are three porphyrin binding enzymes that share a common subunit architecture, while exhibiting highly diverse functionalities (Hofbauer et al., 2021). The subunit architecture is described by two ferredoxin-like folds, which are connected by a flexible linker loop. This loop varies significantly in length and three-dimensional organisation from one enzyme to the other, but also within one enzyme class (Pfanzagl et al., 2018a). Interestingly, catalytically important residues in Clds, ChdCs and DyPs have been identified to be part of this respective loop, linking the N-terminal and the C-terminal domains (Pfanzagl et al., 2018b;Michlits et al., 2020;Schmidt et al., 2021).
Having a simple, robust, adaptable and easily tuneable scaffold, like the one of dimeric α+β barrel proteins, allows to aim at functional diversification and optimization of enzymatic activities for biotechnological applications and processes. Nature has set the tone and proved that this subunit architecture can be used in many different ways, being able to turnover a myriad of different substrates, still with high substrate specificity (Celis and DuBois, 2015). Varying or interchanging this loop in several target enzymes will help to understand evolutional aspects and its specific role for the respective enzyme and also will guide us to potentially novel activities and a better general understanding of structure-function relationships within these family of porphyrin binding dimeric α+β barrel proteins.
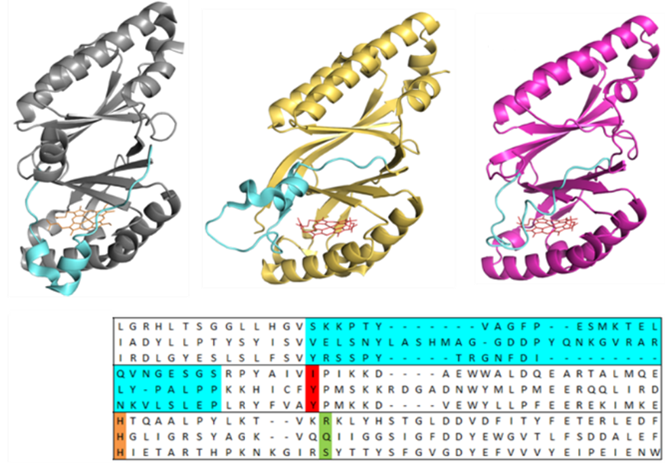
Aims and methods.
ChdC from Corynebacterium diphtheriae and from Listeria monocytogenes, Clds from Nitrospira defluvii and from Cyanothece spp. 7425, and DyP from Klebsiella pneumoniae (always wild-types and loop variants) will be heterologously expressed in E. coli. Further, swapping of the N- and the C-terminal domains, as well as interchanging several domains from all involved enzymes, sampling many combinations, will be assessed in silico by creating AlphaFold models and if the probability to produce a stable protein is high also these artificial enzymes will be produced in vitro, for structural, biophysical and biochemical studies.
Wild-type and mutant proteins will be characterized by a broad set of biochemical/biophysical methods including (i) X-ray crystallography, (ii) high-resolution Cryo-EM studies (in cooperation with Dominika BOREK, UTSouthwestern Medical Center, Dallas, TX, USA), (iii) detailed spectral analysis (UV-vis, electron paramagnetic resonance and resonance Raman spectroscopy) of proteins, and protein-based radicals in different redox- and spin-states (in cooperation with Giulietta SMULEVICH from the Department of Chemistry, University of Florence, Italy), (iv) time-resolved multi-mixing UV-Vis stopped-flow studies in order to analyse the kinetics of interconversion and spectroscopic features of relevant redox intermediates, (v) spectroelectrochemical studies (in cooperation with Gianantonio BATTISTUZZI from the Department of Chemistry, University of Modena and Reggio Emilia, Italy).
Celis, A.I., and Dubois, J.L. (2015). Substrate, product, and cofactor: The extraordinarily flexible relationship between the CDE superfamily and heme. Arch Biochem Biophys 574, 3-17.
Hofbauer, S., Pfanzagl, V., Michlits, H., Schmidt, D., Obinger, C., and Furtmuller, P.G. (2021). Understanding molecular enzymology of porphyrin-binding alpha + beta barrel proteins - One fold, multiple functions. Biochim Biophys Acta Proteins Proteom 1869, 140536.
Michlits, H., Lier, B., Pfanzagl, V., Djinovic-Carugo, K., Furtmüller, P.G., Oostenbrink, C., Obinger, C., and Hofbauer, S. (2020). Actinobacterial coproheme decarboxylases use histidine as distal base to promote Compound I formation. ACS Catal 10, 5405-5413.
Pfanzagl, V., Holcik, L., Maresch, D., Gorgone, G., Michlits, H., Furtmüller, P.G., and Hofbauer, S. (2018a). Coproheme decarboxylases - Phylogenetic prediction versus biochemical experiments. Arch Biochem Biophys 640, 27-36.
Pfanzagl, V., Nys, K., Bellei, M., Michlits, H., Mlynek, G., Battistuzzi, G., Djinovic-Carugo, K., Van Doorslaer, S., Furtmüller, P.G., Hofbauer, S., and Obinger, C. (2018b). Roles of distal aspartate and arginine of B-class dye-decolorizing peroxidase in heterolytic hydrogen peroxide cleavage. J Biol Chem 293, 14823-14838.
Schmidt, D., Serra, I., Mlynek, G., Pfanzagl, V., Hofbauer, S., Furtmuller, P.G., Djinovic-Carugo, K., Van Doorslaer, S., and Obinger, C. (2021). Arresting the Catalytic Arginine in Chlorite Dismutases: Impact on Heme Coordination, Thermal Stability, and Catalysis. Biochemistry 60, 621-634.