
Development of high-throughput-screening of flavin- and heme-dependent oxidoreductases using genetically encoded redox sensors
JOINT-SUPERVISORS: Clemens PETERBAUER, Christian OBINGER
Background.
At VIBT of BOKU two groups work in the field of molecular enzymology and catalysis of oxidoreductases. Flavin-dependent glucose-methanol-choline (GMC) oxidoreductases, a family that includes glucose oxidase, pyranose oxidase, cellobiose dehydrogenase and aryl-alcohol oxidase, are investigated at the Department of Food Science and Technology. These enzymes are implicated in a number of biological processes, and many are interesting for applications in biosensors and biocatalysis (Fraaije et al. 2000). They perform regiospecific oxidations of the CH-OH moiety of various substrates and employ oxygen and/or alternative compounds (quinones, dyes, complexed metal ions) as electron acceptors, with hydrogen peroxide often occurring as principal or secondary by-product. At the Department of Chemistry (Division of Biochemistry) structure-function relationships of heme peroxidases from four superfamilies (Zamocky et al., 2015) are investigated. These enzymes catalyse one- and two-electron oxidation reactions of inorganic and organic compounds thereby consuming hydrogen peroxide. Peroxidases are not only responsible for protection against harmful side-effects of oxygen metabolism in organisms but are also involved in a variety of biosynthetic and degradative functions. As a consequence they are widely used in various biotechnological and chemical transformation reactions.
For flavin- and heme-dependent enzymes numerous engineering efforts are reported, both for scientific reasons as well as towards improved variants for applications. However, the reaction products as well as electron acceptors have low molar mass and are diffusible, and not naturally associated with the enzyme or the cell. Catalytic assays, often using cell-free lysates, are therefore generally performed with artificial compartmentalization in microtiter plates. This requires a lot of lab space, disposable materials and chemicals, as well as time and man-power. Consequently, engineering efforts are often limited to (multiple steps of) saturation mutagenesis (Sygmund et al. 2013, Brugger et al., 2014, Krondorfer et al. 2014). Approaches requiring the screening of very large libraries (CASTing of multiple positions, directed evolution...) are rarely used. Thus, there is clearly a need for high-throughput methodologies that (i) provide a proper association of the encoding gene with the (modified) enzyme and an indicator of the activity, and that allows (ii) the screening of large numbers of clones directly on plate surfaces or cell sorting by FACS in order to explore a substantial sequence space.
Aims and methods.
Genetically encoded redox sensors based on (versions of) the Green Fluorescent Protein (roGFP) have been reported as a powerful tool to measure the intra- and extracellular redox state(s) of cells (Lukyanov and Belousov et al., 2015).
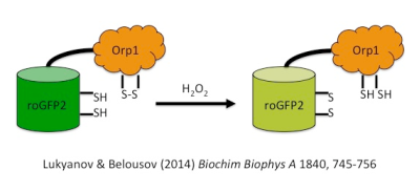
An attractive advancement of this approach is the fusion of redox-sensitive GFPs with thioredoxin-dependent peroxidases that acts as a redox relay: The (non-heme) peroxidase uses H2O2 as oxidant and roGFP as its reducing substrate, if both proteins are fused together. A promising construct is the combination of roGFP2 and the yeast intracellular peroxidase Opr1 resulting in a H2O2-probe with the spectral signatures of roGFP2 (Lukyanov and Belousov 2014).
Workpackage 1: As a starting point of this project conditions for activity screening of representative flavin-dependent enzymes that can be routinely expressed in E. coli (wild-type as well as deletion mutants lacking monofunctional catalase and/or bifunctional catalase-peroxidase) will be established. One model enzyme will be pyranose oxidase (POx, H2O2 producer) from Trametes multicolor. In the beginning wild-type and well characterized mutants will be used for establishment of the roGFP2-Orp1 sensor as a readout for hydrogen peroxide generated by POx. Co-expression of pox and rogfp2-orp1 independently or as a fusion of pox-rogfp2-orp1 for even greater spatial proximity will be tested, and the optimal sequence of cultivation (carbon source, induction and reaction) will be determined. Screening will be tested both on plates with a fluorescent imaging system as well as by FACS.
Workpackage 2: Many flavin- (and heme-) dependent oxidoreductases are secretory proteins. Thus, in addition we aim at adapting the roGFP2-Orp1 redox sensor for extracellular proteins by expressing it in a yeast surface display system, again co-expressed with the also surface-displayed enzyme or as a fusion construct comprising all three proteins to guarantee close proximity. The main challenge in this approach will be the cross-reaction of hydrogen peroxide produced by one enzyme/cell with the redox sensor displayed on neighboring cells. This needs careful exploration of the kinetics of the redox relay and the sustainability of the fluorescence signal to determine optimum reaction conditions. As an alternative, micro-compartmentalization in double emulsions of water-in-oil-in-water (already established for yeast cells) will be employed (Prodanovic et al. 2011).
Workpackage 3: Finally, this screening system will be adapted to screening of H2O2-consuming heme peroxidases and catalases. Here, as a starting point the model enzyme will be the bifunctional catalase-peroxidase (KatG) from Magnaporthe grisea that can easily be expressed intracellularly in an available E. coli strain that lack hydroperoxidases. Many well characterized (bifunctional and monofunctional) KatG variants are available (Gasselhuber et al., 2015) for establishment of optimum screening conditions. It was already demonstrated that intracellular expressed roGFP2-Orp1 responds to extracellular H2O2 concentration in the low micromolar range (Gutscher et al. 2009) allowing to monitor the activity of H2O2 consumption and corresponding disulfide bond transfer via roGFP2. Thus expression of the roGFP2-Orp1 sensor and provision of a constant, low hydrogen peroxide concentration from an external source should result in a constant signal from roGFP2. Conditional co-expression of a hydrogen peroxide-consuming peroxidase would then reduce intracellular hydrogen peroxide levels and result in a drop in signal strength. As a proof of concept the restoration of bifunctional (including catalatic)activity by directed evolution of monofunctional or inactive KatG variants or domains thereof will be tested. The assay should be usable for any H2O2-scavenging or -degrading peroxidase that can be functionally expressed intracellularly in E. coli or yeast.
Brugger, D., Shelswell, C., Huber-Dittes, B., Haltrich, D., Peterbauer, C.K. (2014) Engineering Pyranose 2-Oxidase for Modified Oxygen Reactivity. PLoS ONE 9, 0109242
Fraaije, M., Mattevi, A. (2000) Flavoenzymes: Diverse catalysts with recurrent features. Trends Biochem. Sci. 25, 126-132
Gasselhuber, B., Jakopitsch, C., Zamocky, M., Furtmueller, P.G., Obinger, C. (2015) Mechanistic and functional aspects of catalase-peroxidase. In: Heme Peroxidases (Raven, E., Dunford, H. B., Eds.). Royal Chemical Society (RSC), in press
Gutscher, M., Sobotta, M.C., Wabnitz, G.H., Ballikaya, S., Meyer, A.J., Samstag, Y. and Dick, T.P. (2009) Proximity-based protein thiol oxidation by H2O2-scavenging peroxidases. J. Biol. Chem. 284, 31532-31540
Krondorfer, I., Lipp, K., Brugger, D., Staudigl, P., Sygmund, C., Haltrich, D., Peterbauer, C.K. (2014) Engineering of pyranose dehydrogenase for increased oxygen reactivity. PLoS ONE 9, e91145
Lukyanov, K. and Belousov, V. (2014) Genetically encoded fluorescent redox sensors. Biochim. Biophys. Acta 1840, 745-756
Prodanovic, R., Ostafe, R., Scacioc, A. and Schwanberg, U. (2011) Ultrahigh Throughput Screening system for directed glucose oxidase evolution in yeast cells. Comb. Chem. High Troughp. Scr. 14, 55-60
Sygmund, C., Santner, P., Krondorfer, I., Peterbauer, C.K., Alcalde, M., Nyanhongo, G.S., Guebitz, G.M., Ludwig, R. (2013) Semi-rational engineering of cellobiose dehydrogenase for improved hydrogen peroxide production. Microb. Cell Fact . 12, 38
Zamocky, M., Hofbauer, S., Schaffner, I., Gasselhuber, B., Nicolussi, A., Soudi, M., Pirker, K. F., Furtmueller, P. G., Obinger, C. (2015) Independent evolution of four heme peroxidase superfamilies. Arch. Biochem. Biophys., in press