
Molecular basis of pyruvylated cell wall polymer – SLH-protein interactions
SUPERVISOR: CHRISTINA SCHÄFFER
Background.
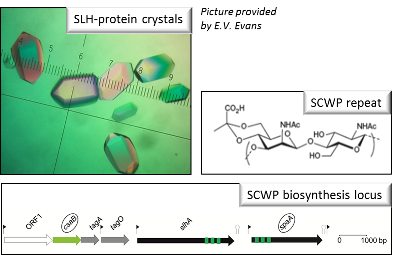
Various Gram-positive bacteria employ S-layer homology (SLH) domain-mediated sorting of cell surface proteins and engage pyruvylated secondary cell wall polymer (SCWP) as a cell wall ligand, with ketal pyruvate linked to β-ManNAc being regarded as an indispensable and ancestral epitope in this binding interaction (Cava et al., 2004; Mesnage et al., 2000). While the backbone structures of pyruvylated SCWPs are similar, revealing a minimum disaccharide [→3)-β-D-ManpNAc-(1→4)-β-D-GlcpNAc-(1→] repeat, non-stoichio-metric modifications contribute to the heterogeneity of some SCWPs and pyruvate ketal was found to be displayed either as a single epitope (e.g., Bacillus anthracis, (Forsberg et al., 2012)) or in multivalent fashion (e.g., Paenibacillus alvei, (Messner et al., 2009)).
The P. alvei SCWP consists of approximately ten disaccharide repeats with each D-ManNAc residue modified with a 4,6-linked pyruvate ketal. We hypothesize that distinct structural features of the exposed pyruvate ketal and amino sugar residues of the SCWP backbone are crucial for the interaction with SLH domains of proteins. To unravel the molecular basis of this binding interaction, the stoichiometrically defined, structurally simple SCWP of P. alvei shall be employed in combination with the S-layer protein SpaA as a cognate, SLH domain-containing binding partner with the three binding motifs TRAE (in SLH1), TVEE (in SLH2), and TRAQ (in SLH3) (Janesch et al., 2013). First crystals of the SpaASLH protein (residues 22-193) have been already obtained (R. Blacker, A. Lopez-Guzman, P. Messner, P. Kosma, C. Sch äffer, S. Evans, manuscript in preparation), providing the basis for the planned co-crystallization with SCWP. The solved structure of unliganded SpaASLH revealed an overall fold similar to B. anthracis SapSLH (25 percent sequence identity), with the most significant structural differences found in the loops connecting the helices. In both structures, there are three lobes with the SCWP-binding residues lining the grooves.
Aims and methods.
By employing a bottom-up approach involving a portfolio of synthetic SCWP fragments (provided by Paul KOSMA) and truncated, soluble and also mutated SpaA, a biological interaction unit shall be determined, characterized, and analyzed in a co-crystal-lized state. To determine the structural interaction basis between SCWP and the SLH pseu-dotrimer, biophysical SpaASLH-binding measurements will be performed. The contribution of the ionic interaction exerted by the carboxylic group of the pyruvate residues will be compared to binding interactions with synthetic carboxyl-reduced analogues. In addition, the individual contributions of the three binding motifs in the SCWP/SpaASLH interaction will be assessed. This study will unravel, if the interprong groove of the SLH pseudotrimer preferentially accommodates SCWP of a certain length and composition, if multiple pyruvate residues are involved in the binding interaction between SCWP and SpaASLH, and if the three binding motifs are equally critical for the interaction. Further, the pyruvyltransferase involved in biosynthesis of P. alvei SCWP will be investigated as a key enzyme for production of biofunctional SCWP.
Experimental set-up:
(i) Recombinant production of rational SpaA protein truncations and mutants (SpaASLH) in E.coli, including a cleavable purification tag. Initial mutants will concern the TVEE and TRAQ motifs to be replaced by TRAE (see below) as well as a rarely discussed conserved motif GIIxG at the N-terminal region of SpaA.
(ii) Biophysical measurements of SpaASLH-SCWP-fragment interactions. To characterize a possible conformational changes of SpaASLH upon binding of the different synthetic SCWP fragments, CD measurements will be performed. To determine the binding affinity and stoich-iometry of the SpaASLH/SCWP interaction, ITC will be employed. The obtained overall binding constants will inform about the tightness of the formed complex. The binding strength bet-ween SpaASLH and the provided SCWP ligands will be determined in SPR experiments. Previously, it was determined that the motifs TRAE, TVEE and TRAQ contribute to the binding of SpaA to varying extents (Janesch et al., 2013). Since in that study, cell wall frag-ments including native SCWP bound to PG was used as a ligand for full-size SpaA carrying mutated binding motifs, the densitometrically quantified complex formation did not exclusively record the interaction of SpaA with the SCWP but also that with the PG. To specifically measure the interaction with SCWP, here, a set of SpaASLH constructs with mutated binding motifs will be used for biophysical binding measurements with synthetic SCWP ligands. (iii) Depending on the progress of the work, data will be complemented by SCWP-fragment/ protein docking studies. (iv) To identify a smallest biological interaction unit, structure determination by X-ray crystal-lography will be performed utilizing an already confined set of possible interaction partners according to the biophysical measurements (ii). Based on the findings that TRAE is a frequent binding motif in SLH-proteins and that it has, according to our in vitro binding assay (Janesch et al., 2013), the most pronounced influence of the three SpaA binding motifs on the interaction with SCWP it will be investigated, if the provision of TRAE motifs in all three SLH domains would favor the interaction.
(iii) Insight into P. alvei SCWP pyruvylation. Recombinant production of the P. alvei pyruvyltransferase CsaB and set-up of an in vitro pyruvylation assay. By using synthetic SCWP fragments of different lengths as pyruvylation targets, it will be unraveled at which stage of polymer formation pyruvylation of ManNAc occurs.
This study will lead to a better understanding of how bacteria stick their cell wall together, pinpointing potential strategies of how to interfere with the bacterial cell envelope architecture to control the biological status of bacteria. It might well be that the investigated binding mechanism is more prevalent than currently anticipated, since SCWPs might escape from detection of pyruvate due to the acid treatment frequently employed for SCWP release from bacterial cell walls.
Collaborations within this thesis will include KOSMA (organic synthesis of SCWP fragments), OBINGER (CD, ITC) and OOSTENBRINK (SCWP-fragment/protein docking studies). International cooperations include EVANS (X-ray crystallography).
Cava, F., de Pedro, M. A., Schwarz, H., Henne, A., Berenguer, J. (2004) Binding to pyruvylated compounds as an ancestral mechanism to anchor the outer envelope in primitive bacteria. Molecular microbiology 52, 677-690.
Forsberg, L. S., Abshire, T. G., Friedlander, A., Quinn, C. P., Kannenberg, E. L., Carlson, R. W. (2012) Localization and structural analysis of a conserved pyruvylated epitope in Bacillus anthracis secondary cell wall polysaccharides and characterization of the galactose-deficient wall polysaccharide from avirulent B. anthracis CDC 684. Glycobiology 22, 1103-1117.
Janesch, B., Messner, P., Schäffer, C. (2013) Are the surface layer homology domains essential for cell surface display and glycosylation of the S-layer protein from Paenibacillus alvei CCM 2051T? Journal of bacteriology 195, 565-575.
Mesnage, S., Fontaine, T., Mignot, T., Delepierre, M., Mock, M., Fouet, A. (2000) Bacterial SLH domain proteins are non-covalently anchored to the cell surface via a conserved mechanism involving wall polysaccharide pyruvylation. The EMBO journal 19, 4473-4484.
Messner, P., Egelseer, E. M., Sleytr, U. B., Schäffer, C. (2009) Bacterial surface layer glycoproteins and "non-classical" secondary cell wall polymers. in: A P Moran, P J Brennan, O Holst, and M von Itzstein (ed), Microbial glycobiology: structures, relevance and applications, 109-128.