
O-glycan engineering of N. benthamiana for structure-function characterization of human IgA1
SUPERVISORS: Eva STÖGER and Richard STRASSER
Background.
N- and O-glycans of proteins play central roles in many biological processes. Key to the understanding of glycan function are systems that allow the controlled expression of glycoproteins with desired glycan modifications. Although the N-glycan processing pathway of different expression hosts has been successfully modified in the past, comparatively little attention has been paid to the generation of customized O-linked glycans.
Mucin-type O-glycosylation is the most abundant form occurring on secretory mammalian proteins. Mucin-type O-glycosylation is initiated in the Golgi by a family of 20 polypeptide GalNAc-transferase isoenzymes (GalNAc-Ts) that attach GalNAc to the hydroxyl groups of serine or threonine in proteins (GalNAc-Ser/Thr linkage). After the initiation, the sugar chains are extended in a stepwise manner by incorporation of galactose, sialic acids and other sugar residues.
Plants lack mucin-type O-glycans, as they contain no endogenous GalNAc-Ts. Consequently, plants are an attractive system for de novo engineering of the required glycosylation steps to produce customized O-glycans in a controlled manner (hardly achievable in mammalian cells). Despite this advantage, a major drawback of plants is the hydroxylation and subsequent plant-specific glycosylation of proline residues that are frequently found next to O-glycosylation sites. The presence of these non-human posttranslational modifications has been described for human erythropoietin (EPO) and immunoglobulin A1 (IgA1) expressed in plants (Castilho et al., 2012; Göritzer et al., 2017). Co-expression of human GalNAc-Ts leads to the incorporation of GalNAc residues on plant-produced recombinant glycoproteins, but the hydroxyproline (Hyp) residues are abundant resulting in glycoproteins with very heterogeneous plant-specific and mammalian-type modifications. As a consequence, the role of defined mucin-type O-glycans cannot be examined on these glycoproteins.
IgAs are an abundant antibody class that mediate immune protection at mucosal surfaces or in plasma and recombinant IgAs are promising biopharmaceuticals for treatment of infections and cancer. Compared to IgGs and other IgA isotypes, human IgA1 has an extended hinge region (16 amino acids, rich in Pro, Ser and Thr) that is commonly decorated with 3 to 6 mucin-type O-glycans. The heavily O-glycosylated IgA1 hinge region likely affects the conformational variability of IgA1 by making it less flexible. The specific IgA1 hinge region contributes to an extended T-shaped IgA1 structure that appears important for the biological activity since a greater reach between its two antigen-binding sites can be obtained, allowing this immunoglobulin to achieve high-avidity interaction with more distantly spaced antigens (Böhm et al., 1999). Fca-receptor binding at the IgA1 Fca CH2-CH3 junction causes conformational changes at the hinge region and may induce long-range conformational changes in the antigen binding region (Posgai et al., 2018). The role of the O-glycans in this process is unclear.
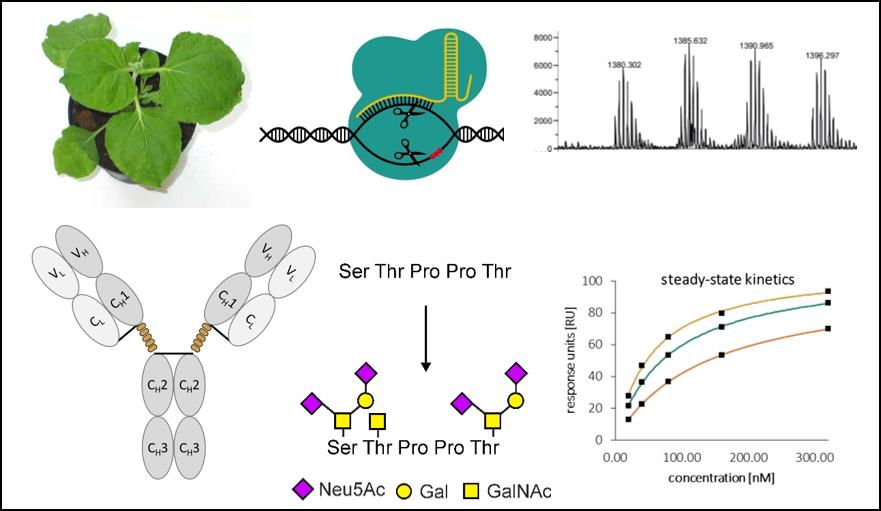
To assess the impact of O-glycosylation on structure-function relationships of IgA1, a suitable expression platform is needed which allows the production of IgA1 variants bearing defined O-glycans. The tobacco-related species Nicotiana benthamiana has emerged as promising host for expression of recombinant glycoproteins with tailored glycans (Castilho et al., 2012; Göritzer et al., 2017). Together with Friedrich ALTMANN we have recently identified three N. benthamiana prolyl-4-hydroxylases (P4Hs) involved in Hyp-formation of recombinant plant-produced IgA1 (unpublished). These identified P4Hs provide targets for CRISPR/Cas9-mediated genome engineering to generate a N. benthamiana platform lacking the interfering P4H activities. Consequently, we hypothesize that functional IgA1 variants lacking plant-specific O-glycans and furnished with customized mucin-type O-glycans can be generated by extensive glyco-engineering of N. benthamiana. P4H knockout plants will be generated by multiplex genome-editing and the missing mammalian glycosyltransferases (GalNAc-Ts) and metabolic pathways (e.g. for CMP-sialic acid biosynthesis) will be introduced by transient expression. Recombinant IgA1 with defined O-glycans will be characterized by biochemical/physical techniques and functional assays. The PhD student will be supervised by Richard STRASSER (O-glycan engineering, IgA1 expression) and Eva STÖGER (advanced genome editing and plant transformation). In addition, there will be close cooperation with Friedrich ALTMANN (mass spectrometry, characterization of P4H candidates), Christian OBINGER (spectroscopy, calorimetry) and Rupert TSCHELIESSNIG (small-angle light scattering).
Aims and Methods.
This project aims to establish a robust expression platform for recombinant glycoproteins with tailored O-glycans in order to analyse the impact of O-glycosylation on structure, conformational and thermal stability as well as functional properties (e.g. IgA1 antigen binding and interaction with host receptors).
For each of the three identified P4H candidates involved in IgA1 Hyp formation both alleles (at least six genes – allotetraploid plant) will be inactivated using multiplex CRISPR/Cas9 genome editing (Kapusi et al., 2017; Jansig et al., 2019). The target gene sequences will be verified and screened for regions of homology that can be targeted by the same guide RNAs (gRNAs). Candidate gRNAs will be evaluated for potential off‐targets, and in each experiment more than one target sgRNA per gene will be applied to trigger segment deletions facilitating efficient screening by PCR (Kapusi and Stoger, 2018). Multiplexing will be achieved via Cas6 (Csy4)-mediated precise processing of polycistronic mRNA transcripts driven by a strong PolII promoter. If necessary, doubled haploid pants will be generated after crossing individual genome edited plants, allowing the recovery of true breeding lines that are instantly homozygous for the edited alleles (Kapusi and Stoger, 2018). Knockouts will be confirmed by PCR, DNA sequencing and mass spectrometry. Monomeric HER2-binding IgA1 (Göritzer et al., 2017) will be transiently expressed in genome edited N. benthamiana with or without the mammalian O-glycan biosynthesis pathway using transient expression vectors (Castilho et al., 2012). The recombinant IgA1 glycoforms will be purified and the IgA1 hinge region will be subjected to LC-ESI-MS analysis to determine the O-glycan composition (Göritzer et al., 2017).
The variants with defined O-glycans will be investigated for their overall structure and homogeneity [size exclusion chromatography combined with multi-angle light scattering, circular dichroism (CD) spectroscopy], proteolytic susceptibility in the hinge region as well as conformational and thermal stability in the presence or absence of the antigen and/or Fca-receptor (CD and fluorescence spectroscopy, differential scanning calorimetry) (Göritzer et al., 2017). Small-angle X-ray scattering (SAXS) will be employed for evaluation of the O-glycan structure-function relationships.
Taken together, our study aims to establish a robust plant-based expression system for IgA1 and other O-glycosylated recombinant proteins in order to systematically evaluate the impact of defined O-glycans on protein function.
Boehm MK, Woof JM, Kerr MA, Perkins SJ. (1999) The Fab and Fc fragments of IgA1 exhibit a different arrangement from that in IgG: a study by X-ray and neutron solution scattering and homology modelling. J. Mol. Biol. 286, 1421-1447. doi: 10.1006/jmbi.1998.2556
Castilho A, Neumann L, Daskalova S, Mason HS, Steinkellner H, Altmann F, Strasser R. (2012) Engineering of sialylated mucin-type O-glycosylation in plants. J. Biol. Chem. 287, 36518-36526. doi: 10.1074/jbc.M112.402685
Göritzer K, Maresch D, Altmann F, Obinger C, Strasser R. (2017) Exploring Site-Specific N-Glycosylation of HEK293 and Plant-Produced Human IgA Isotypes. J. Proteome Res. 16, 2560-2570. doi: 10.1021/acs.jproteome.7b00121
Jansing J, Sack M, Augustine SM, Fischer R, Bortesi L. (2019) CRISPR/Cas9-mediated knockout of six glycosyltransferase genes in Nicotiana benthamiana for the production of recombinant proteins lacking β-1,2-xylose and core α-1,3-fucose. Plant Biotechnol J. 2019 17, 350-361. doi: 10.1111/pbi.12981
Kapusi E, Corcuera-Gómez M, Melnik S, Stoger E. (2017) Heritable Genomic Fragment Deletions and Small Indels in the Putative ENGase Gene Induced by CRISPR/Cas9 in Barley. Front. Plant Sci. 8, 540. doi: 10.3389/fpls.2017.00540
Kapusi E and Stoger E. (2018) Detection of CRISPR/Cas9-Induced Genomic Fragment Deletions in Barley and Generation of Homozygous Edited Lines via Embryogenic Pollen Culture. Methods Mol Biol. 1789, 9-20. doi: 10.1007/978-1-4939-7856-4_2.
Posgai MT, Tonddast-Navaei S, Jayasinghe M, Ibrahim GM, Stan G, Herr AB. (2018) FcαRI binding at the IgA1 CH2-CH3 interface induces long-range conformational changes that are transmitted to the hinge region. Proc. Natl. Acad. Sci. USA. 115, E8882-E8891. doi: 10.1073/pnas.1807478115