
Effects of N-glycans on structure and dynamics of IgAs and their implications on receptor binding
JOINT-SUPERVISORS: Richard Strasser and Rupert Tscheließnig
Background.
Immunoglobulin A (IgA) is the most abundant antibody produced by humans. IgAs mediate immune protection at mucosal surfaces or in plasma and recombinant IgAs are potent biopharmaceuticals for treatment of infections and cancer (Macpherson et al., 2018; Brandsma et al., 2019). Compared to IgG-based drugs, the potential of recombinant IgAs as therapeutic antibodies is not fully explored and the structure-function relationships remain poorly understood. The conserved N-glycan in the Fc domain of IgGs plays a crucial role for the IgG structure and modulates the binding to specific Fc-receptors (Yamaguchi and Barb, 2019; Wang & Ravetch, 2019). By contrast, comparatively little is known how N-glycans impact the protein structure and function of IgAs. The two human IgA isotypes (IgA1 and IgA2) carry two to five N-glycans on the heavy chain. While in IgG1 and other immunoglobulins, the N-glycan in the CH2 domain is confined within the interstitial space between two heavy chains, the conserved CH2 domain N-glycans of IgAs are located at the external surface of the protein (Mattu et al., 1998). Compared to the IgA-Fca-receptor interaction, IgGs display a different binding mode to Fc-receptors (Herr et al., 2003). These distinct structural properties lend the IgA antibody class a unique array of functional capabilities.
Our group has established Nicotiana benthamiana and human HEK293F cell-based expression systems. These allow efficient generation of monomeric as well as dimeric IgA1 and IgA2 variants with defined N-glycans (Göritzer et al., 2017). Our data indicate site-specific processing of N-glycans in the CH2 region, which is likely caused by protein-glycan interactions. Subsequent Fca-receptor binding studies revealed that the N-glycans on monomeric IgA variants do not play a major role in the IgA-Fca-receptor interaction (Göritzer et al., 2019), whereas the N-glycans of the Fca-receptor affect binding to IgA.
Aims and methods.
We will analyse the impact of N-glycosylation on the structure-function relationships of different IgA formats and elucidate the conformation of IgA glycoforms in solution and bound to receptors by small angle X-ray scattering (SAXS). From pair density distributions we will molecular model structures that help to explain the role of N-glycans for IgAs, for IgA-receptor interactions and for IgA-antigen-receptor interactions. The IgA-Fca-receptor interaction is well studied (Mattu et al., 1998; Herr et al., 2003; Göritzer et al., 2019) and will serve as benchmark for our proposed approaches. The binding mode of the less characterized IgA receptor Fc receptor-like 4 (FcRL4) (Wilson et al., 2012) will then be studied in detail by the prospective PhD student. FcRL4 is the only known inhibitory IgA receptor with a suggested role in the regulation of the B cell response. Like the Fca-receptor, it is a potential target for therapeutics e.g. to combat chronic viral infections. The binding mode of this receptor and a putative role of N-glycans are currently unknown. We will monitor IgA glycoforms and their role for the FcRL4-receptor interaction in solution by SAXS. The data will be evaluated by molecular modelling. We will use Reverse Monte Carlo (RMC) and Molecular Dynamics Simulations (MDS). Comparable approaches elucidated the conformational characteristics of different IgG Fc domains (Remesh et al., 2018), glycosylated and aglycosylated IgG Fc regions (Yageta et al., 2019) and differentially O-glycosylated IgAs (Hui et al., 2015), already.
We pose a set of research questions: Do the IgA CH2 N-glycans have an impact on the protein conformation in solution? How does the FcRL4-receptor bind to IgA variants and IgA immune complexes? Is the FcRL4-receptor binding dependent on IgA N-glycans? Do FcRL4-receptor N-glycans play a role for IgA binding?
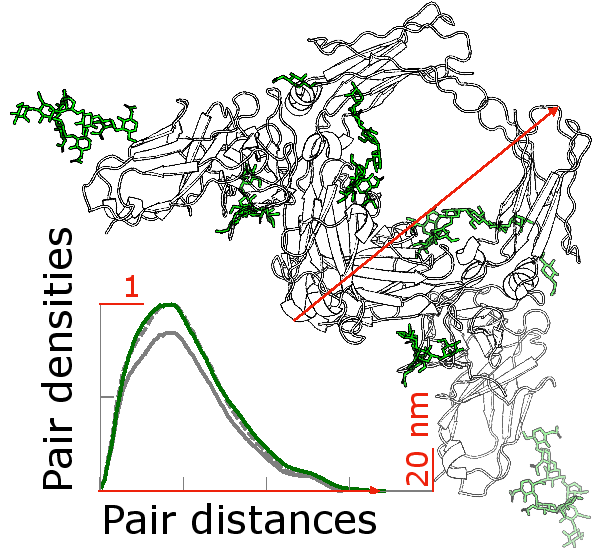
Fig. 1.Glycosylated (green) and non glycosylated (grey) human FcaRI bound to IgA1-Fc: The differences in the glycosylation pattern of proteins can be characterised by the pair density distributions. Different pair density distributions were computed: in green, they correspond to the glycosylated and in grey to the non glycosylated model. These pair density distributions are experimental quantities that can be assessed by scattering experiments. Through comparison of theoretical and experimental pair density distributions, the second will be determined by molecular simulations, the prospective PhD student will elucidate the structure-function relationship of the glycosylation pattern of proteins.
To produce IgA variants lacking the N-glycans in the CH2 domain the consensus N-glycosylation sites (1 site in IgA1 and 2 sites in IgA2) will be mutated using site-directed mutagenesis. Monomeric HER2-binding IgA1 and IgA2 (Göritzer et al., 2017) as well as the newly generated mutant variants will be transiently expressed in glyco-engineered N. benthamiana to produce IgAs with homogeneous human-type complex N-glycans. The recombinant IgAs will be purified by affinity and size exclusion chromatography and subjected to LC-ESI-MS analysis to determine the site-specific N-glycan composition (Göritzer et al., 2017). HER2 antigen binding will be determined by ELISA and the thermal stability in the presence or absence of the HER2 antigen will be measured by differential scanning calorimetry (Göritzer et al., 2017). SAXS will be employed for evaluation of the N-glycan structure-function relationships in solution. The extracellular domain of the human FcRL4-receptor will be produced in human HEK293F cells as described previously for the Fca-receptor (Göritzer et al., 2019). The purified receptor will be used for in vitro binding studies with IgA variants using SPR spectroscopy and isothermal titration calorimetry (ITC) and for the determination of the in solution structure using SAXS in the presence of IgA variants and IgA-HER2 immune complexes. The PhD student will be jointly supervised by Richard STRASSER (IgA and receptor expression, purification and glyco-engineering) and Rupert TSCHELIESSNIG (SAXS and biophysical characterization). Additional internal collaboration partners are Friedrich ALTMANN (mass spectrometry, glycan analysis) and Chris OOSTENBRINK (molecular modelling).
REFERENCES:
Brandsma AM, Bondza S, Evers M, Koutstaal R, Nederend M, Jansen, JHM, Rösner T, Valerius T, Leusen JHW and Ten Broeke T (2019) Potent Fc receptor signaling by IgA leads to superior killing of cancer cells by neutrophils compared to IgG. Front. Immunol. 10: 704 doi: 10.3389/fimmu.2019.00704.
Göritzer K, Maresch D, Altmann F, Obinger C, Strasser R (2017) Exploring Site-Specific N-Glycosylation of HEK293 and Plant-Produced Human IgA Isotypes. J. Proteome Res. 16: 2560-2570. doi: 10.1021/acs.jproteome.7b00121
Göritzer K, Turupcu A, Maresch D, Novak J, Altmann F, Oostenbrink C, Obinger C, Strasser R (2019) Distinct Fc alpha receptor N-glycans modulate the binding affinity to immunoglobulin A (IgA) antibodies. J Biol Chem. 294: 13995-14008. doi: 10.1074/jbc.RA119.009954
Herr AB, Ballister ER, Bjorkman PJ (2003) Insights into IgA-mediated immune responses from the crystal structures of human FcalphaRI and its complex with IgA1-Fc. Nature 423: 614-620. doi: 10.1038/nature01685
Hui GK, Wright DW, Vennard OL, Rayner LE, Pang M, Yeo SC, Gor J, Molyneux K, Barratt J, Perkins SJ. (2015) The solution structures of native and patient monomeric human IgA1 reveal asymmetric extended structures: implications for function and IgAN disease. Biochem J. 471: 167-185. doi: 10.1042/BJ20150612
Macpherson AJ, Yilmaz B, Limenitakis JP, Ganal-Vonarburg SC (2018) IgA function in relation to the intestinal microbiota. Annu Rev Immunol. 36: 359–381. doi: 10.1146/annurev-immunol-042617-053238
Mattu TS1, Pleass RJ, Willis AC, Kilian M, Wormald MR, Lellouch AC, Rudd PM, Woof JM, Dwek RA (1998) The glycosylation and structure of human serum IgA1, Fab, and Fc regions and the role of N-glycosylation on Fcα receptor interactions. J Biol Chem. 273: 2260-2272. doi: 10.1074/jbc.273.4.2260
Remesh SG, Armstrong AA, Mahan AD, Luo J, Hammel M (2018) Conformational Plasticity of the Immunoglobulin Fc Domain in Solution. Structure 26: 1007-1014 doi: 10.1016/j.str.2018.03.017
Wang TT, Ravetch JV (2019) Functional diversification of IgGs through Fc glycosylation. J Clin Invest. 129: 3492-3498. doi: 10.1172/JCI130029
Wilson TJ, Fuchs A, Colonna M (2012) Cutting edge: human FcRL4 and FcRL5 are receptors for IgA and IgG. J Immunol 188: 4741-4745. doi: 10.4049/jimmunol.1102651
Yageta S, Imamura H, Shibuya R, Honda S. (2019) CH2 domain orientation of human immunoglobulin G in solution: Structural comparison of glycosylated and aglycosylated Fc regions using small-angle X-ray scattering. Mabs 11: 453-462. doi: 10.1080/19420862.2018.1546086
Yamaguchi, Y, Barb AW (2019) A synopsis of recent developments defining how N-glycosylation impacts immunoglobulin G structure and function Glycobiology published: 06 September 2019. doi: 10.1093/glycob/cwz068