
Targeting interactions of enzymes on cell wall polymers
SUPERVISOR: Roland LUDWIG
Background.
Many enzymes feature substrate binding domains that are not part of their active site and are often located on a different domain. Cellulosic and hemicellulosic enzymes use carbohydrate binding modules (CBM) of which 101 families are currently classified in the CAZy database. CBMs bind to carbohydrate polymers and establish an equilibrium of bound and soluble enzymes. Only substrate-bound enzymes are catalytically active and therefore the binding strength of the CBM governs the activity by the number of bound enzymes and their dynamic distribution over the polymer surface. However, only soluble enzymes can target accessible, newly cleared sites in the cell wall’s polymer network and thereby propagate enzymatic action. In the case of lytic polysaccharide monooxygenase (LPMO) about half of all sequenced genes have a C-terminal CBM. To date, it is unknown why some LPMOs with the same substrate specificity differ in this regard (Støpamo, 2024). In addition, direct interaction with its auxiliary enzyme cellobiose dehydrogenase (CDH) is necessary for catalysis. The activity of LPMO is therefore not only regulated by a CBM, but also by a weak, transitional protein-protein interaction to a CDH, which also can have a CBM (Kracher, 2016). The evolutionary role of the CBM for the function of these two enzymes, the CBM’s necessity for catalytic substrate turnover by binding to the substrate surface, or its targeting function towards accessible substrate surfaces in the plant cell wall has not been elucidated yet.
Hypotheses.
1. The C-terminal CBM on LPMO has an optimal affinity to achieve the highest enzymatic activity, which is not too low or too high but in a defined Goldilocks zone. 2. The absence of a CBM or a CBM with a low affinity increases the soluble fraction of LPMO and increases the targeting of newly accessible substrate sites in the plant cell wall.
Methods.
Two fungal model enzymes binding to cellulose and to each other are investigated by the proposed project: LPMO and CDH. LPMO and CDH from Neurospora crassa with and without a CBM, as well as with a CBM of engineered affinity will be recombinantly produced in Komagataella phaffii and purified by column chromatography (Sygmund, 2012). CBMs will be tuned by protein engineering for lower substrate affinity, if a higher affinity is needed, a combination of two C-terminal CBMs will be fused to the enzyme. The PhD candidate will investigate different methods and strategies for the molecular modelling, simulation and rational engineering of the CBMs. Additionally, the CDH cytochrome domain will be modified for a lower and a higher affinity towards LPMO. The PhD candidate will also select and establish methods to study the binding, activity and distribution of LPMO and CDH on cellulose as a defined substrate and in the plant cell wall under near natural conditions. To determine the kinetics of substrate binding on cellulose a kinetic assay (e.g., a turbidimetric or fluorometric method) will be established (Filandr, 2020). The kinetics of enzyme mobility and substrate targeting in the cell wall will be studied by confocal microscopy of fluorescence-tagged enzymes (Gacias-Amengual, 2022).
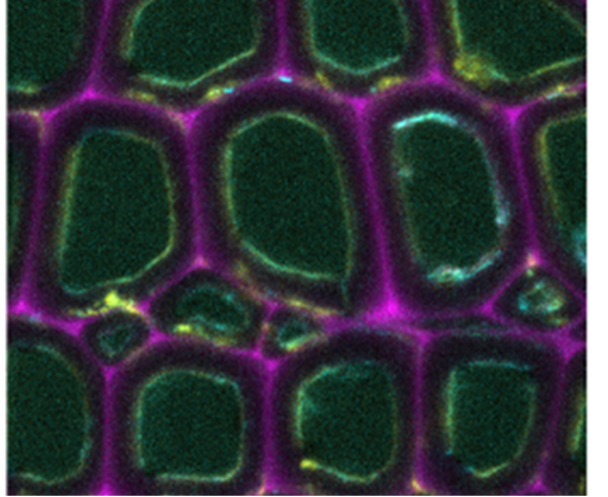
Surface plasmon resonance for affinity measurements will be supported by molecular simulations. The targeting of plant wall polymers by unmodified or engineered enzymes will be studied in situ by scanning electrochemical microscopy (Chang, 2022). The interaction of LPMO and CDH and the resulting electron transfer rate will be evaluated by fast kinetic biochemical and electrochemical methods.
REFERENCES
1. Støpamo, F. G., I. Sulaeva, D. Budischowsky, J. Rahikainen, K. Marjamaa, A. Potthast, K. Kruus, V. G. H. Eijsink and A. Várnai (2024) Oxidation of cellulose fibers using LPMOs with varying allomorphic substrate preferences, oxidative regioselectivities, and domain structures. Carbohydrate Polymers 330, 121816. doi:
10.1016/j.carbpol.2024.121816
2. Kracher, D., S. Scheiblbrandner, A. K. Felice, E. Breslmayr, M. Preims, K. Ludwicka, D. Haltrich, V. G. Eijsink and R. Ludwig (2016) Extracellular electron transfer systems fuel cellulose oxidative degradation. Science 352, 1098-1101. doi: 10.1126/science.aaf3165
3. Sygmund, C., D. Kracher, S. Scheiblbrandner, K. Zahma, K. G. Felice Alfons, W. Harreither, R. Kittl and R.
Ludwig (2012) Characterization of the two Neurospora crassa cellobiose dehydrogenases and their connection to oxidative cellulose degradation. Applied and Environmental Microbiology 78, 6161-6171. doi: 10.1128/AEM.01503-12
4. Filandr, F., P. Man, P. Halada, H. Chang, R. Ludwig and D. Kracher (2020) The H2Osub>2-dependent activity of a fungal lytic polysaccharide monooxygenase investigated with a turbidimetric assay. Biotechnol Biofuels 13, 37. doi: 10.1186/s13068-020-01673-4
5. Gacias-Amengual, N., L. Wohlschlager, F. Csarman and R. Ludwig (2022) Fluorescent Imaging of Extracellular Fungal Enzymes Bound onto Plant Cell Walls. Int J Mol Sci 23. doi: 10.3390/ijms23095216
6. Chang, H., N. Gacias Amengual, A. Botz, L. Schwaiger, D. Kracher, S. Scheiblbrandner, F. Csarman and R. Ludwig (2022) Investigating lytic polysaccharide monooxygenase-assisted wood cell wall degradation with microsensors. Nat Commun 13, 6258. doi: 10.1038/s41467-022-33963-w