
Who does what – and where? Diverse roles of bacterial peroxidases and their location
SUPERVISOR: Clemens PETERBAUER
Background.
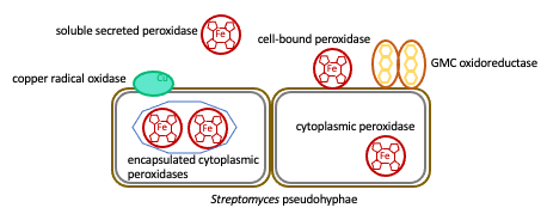
Bacteria like Streptomyces or Amycolatopsis each harbor several genes annotated as (dye-decolorizing) peroxidases or deferrochelatases/peroxidases. Some contain putative signal peptides (with or without cell-anchoring motif), and encapsulin-packaging motifs and downstream-located encapsulin genes are found (Rahmanpour, 2013), suggesting distinct roles. For Streptomyces lividans DtpA and a copper radical oxidase, roles in morphogenesis have been shown (Petrus, 2016), but other roles are discussed (modification of lignin constituents, oxidative stress protection) (Bugg, 2020; Lien, 2021). Whichever function, peroxidase activity requires spatial and/or temporal control for resource efficiency and to avoid cellular damage, and depends on auxiliary activities like enzymes providing hydrogen peroxide or reduced metal ions. As of now, only few of the above-mentioned targeting features are experimentally investigated.
Hypotheses.
1. Bacterial peroxidases have a role in cell physiological processes or in lignin modification and metabolization – or both. 2. Identifying the location of a given enzyme as well as that of auxiliary enzymes required for activity will allow us to differentiate between these major roles.
Approach.
We will express sets of peroxidase-encoding genes from model organisms and determine the subcellular location of the enzymes (cytoplasmic / encapsulated / secretory / surface-attached). Genes encoding enzymes with putative packaging signals will be co-expressed with their downstream located encapsulin gene. Differential labelling systems will be used for tracking of the nascent polypeptides from translation to destination (Hoang, 2023). For secretory enzymes without cell anchor, the affinity to extracellular matrices such as the bacterial or plant cell wall will be determined. Candidate genes for auxiliary enzymes like pyranose oxidase (Virginia, 2023) and copper radical oxidase (Whittaker, 2006) will be co-expressed and possible co-location and interaction will be determined by approaches such as split-luciferase or split-GFP assays. Subsequently, we will generate deletion strains lacking the respective genes and determine the phenotypic consequences to differentiate roles in cell physiological processes or lignin modification and metabolization.
REFERENCES
1. Bugg, T. D. H., J. J. Williamson and G. M. M. Rashid (2020) Bacterial enzymes for lignin depolymerisation: new biocatalysts for generation of renewable chemicals from biomass. Curr Opin Chem Biol 55, 26-33. doi: 10.1016/j.cbpa.2019.11.007
2. Hoang, M. N. and C. Peterbauer (2023) Double-labeling method for visualization and quantification of membrane-associated proteins in Lactococcus lactis. Int J Mol Sci 24. doi: 10.3390/ijms241310586
3. Lien, K. A., K. Dinshaw, R. J. Nichols, C. Cassidy-Amstutz, M. Knight, R. Singh, L. D. Eltis, D. F. Savage and S. A. Stanley (2021) A nanocompartment system contributes to defense against oxidative stress in Mycobacterium tuberculosis. Elife 10. doi: 10.7554/eLife.74358
4. Petrus, M. L., E. Vijgenboom, A. K. Chaplin, J. A. Worrall, G. P. van Wezel and D. Claessen (2016) The DyP-type peroxidase DtpA is a Tat-substrate required for GlxA maturation and morphogenesis in Streptomyces. Open Biol 6, 150149. doi: 10.1098/rsob.150149
5. Rahmanpour, R. and T. D. Bugg (2013) Assembly in vitro of Rhodococcus jostii RHA1 encapsulin and peroxidase DypB to form a nanocompartment. FEBS J 280, 2097-2104. doi: 10.1111/febs.12234
6. Virginia, L. J. and C. Peterbauer (2023) Localization of pyranose 2-oxidase from Kitasatospora aureofaciens: A step closer to elucidate a biological role. Int J Mol Sci 24. doi: 10.3390/ijms24031975
7. Whittaker, M. M. and J. W. Whittaker (2006) Streptomyces coelicolor oxidase (SCO2837p): a new free radical metalloenzyme secreted by Streptomyces coelicolor A3(2). Arch Biochem Biophys 452, 108-118. doi: 10.1016/j.abb.2006.06.020